
Wir befinden uns am Beginn einer neuen Epoche: dem Erdmittelalter, 250 Millionen Jahre vor der Gegenwart. Die Trias beginnt -- sie wird 50 Millionen Jahre dauern.
Die Welt steht nun, am Beginn der Trias, vor einem Neubeginn. Wie schon im Perm zuvor ist fast die gesamte Landmasse zu einem einzigen Superkontinent vereint: Pangäa. Das Klima in Pangäa ist zumeist kontinental, heiß und trocken. Große Wüsten und trockene Hochebenen bedecken den Superkontinent, u.a. das heutige Europa. Sogar die Polargebiete sind relativ warm und (anders als in der Gegenwart) nicht von Eiskappen bedeckt. Zwischen den trockenen Wüsten und den Polargebieten gibt es teilweise größere Waldgebiete.
Viele Lebensräume sind durch das Massensterben am Ende des Perm frei geworden, und die Überlebenden beginnen nun, diese Lebensräume neu zu erobern.
Bei der Rückeroberung der Lebensräume sind manche Arten schneller als andere, und manche alte Vorherrschaft wird gebrochen. Ammoniten drängen die Perlboote ins Abseits und entwickeln eine große Artenvielfalt. Muscheln verdrängen langsam die Armfüßer. Auch die Amphibien erholen sich wieder. Einer der größten Lurche aller Zeiten lebt in der Trias: der mehr als 4 Meter lange Mastodonsaurus (der natürlich -- trotz seines Namens -- als Amphibium kein Saurier ist):

Bei den Landpflanzen setzen sich in trockeneren Gebieten die Samenpflanzen weiter gegen die Sporenpflanzen durch, da letztere auf genügend Feuchtigkeit angewiesen bleiben, die im kontinental-heißen Klima Pangäas selten ist. Erste Blütenpflanzen (Angiospermen) erscheinen gegen Ende der Trias.
Eine besondere Blütezeit erleben in der Trias die Reptilien. Hier eine (unvollständige) Übersicht (genauere Details im Buch):
In der Trias spaltet sich eine Gruppe von den Cynodonten ab, die man als die ersten Säugetiere bezeichnen kann. Ein Vertreter von ihnen ist das Megazostrodon, ein etwa 10 cm langes spitzmausähnliches Tier, das sich vermutlich von Insekten ernährt (siehe dazu z.B. http://www.palaeos.com/Vertebrates/Units/Unit420/420.200.html ).

Ein Modell des Megazostrodons im Natural History Museum, London.
Quelle:
Wikimedia Commons File:Megazostrodon.jpg (Urheber Nordelch),
Quelle dort: http://www.flickr.com/photos/89637257@N00/2052121799/,
in Wikimedia lizenziert unter der Creative Commons-Lizenz
Attribution ShareAlike 2.0.
Im Erdmittelalter spielen die kleinen Säugetiere nur eine geringe Rolle gegenüber den dominierenden Sauriern. So wie die Dinosaurier in Form der Vögel bis in die Gegenwart fortbestehen, so besteht die urtümliche Reptiliengruppe der Cynodonten in Form der Säugetiere bis in die Gegenwart fort.

Krokodil, Quelle:
Wikipedia Commons File:China-Alligator.jpg,
Public domain picture from U.S. Fish and Wildlife Service,
Link

Chuckwalla (Sauromalus obesus) und Halsbandleguan (Crotaphytus collaris),
Quelle:
Wikimedia Commons File:Bristol.zoo.collared.and.western.chuckwallah.arp.jpg,
vom Autor Adrian Pingstone (2005) dort freigegeben (Public Domain).

Brückenechse, Quelle:
Wikipedia Commons File:Tuatara.jpg,
demnach gemeinfrei (Public Domain).

Schildkröten,
Quelle: Wikipedia Commons File:Haeckel Chelonia.jpg,
aus Ernst Haeckel's Kunstformen der Natur (1904), copyright demnach abgelaufen.
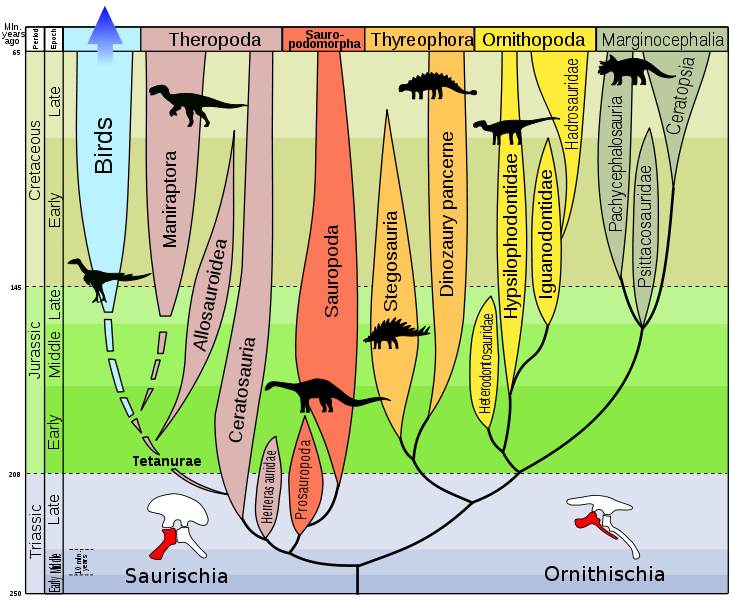
Die Evolution der Dinosaurier. Links oben sind bereits die Vögel (birds) zu erkennen.
Quelle:
Wikimedia Commons File:Evolution of dinosaurs by Zureks.svg von Wikimedia-User Zureks,
dort lizensiert unter der
Creative Commons Attribution ShareAlike 3.0 License.
Die ersten Dinosaurier am Anfang der Trias sind noch recht klein.
Zu den ältesten Dinosauriern gehören
die fleischfressenden Theropoden Herrerasaurus
und Coelophysis
(entsprechende Fossilienfunde sind etwa 225 Millionen Jahre alt).

Herrerasaurus.
Quelle:
Wikimedia Commons File:Herrerasaurus BW.jpg von Nobu Tamura,
dort lizensiert unter
Creative Commons
Attribution 2.5 License.
In der späteren Trias entwickeln sich dann auch große Dinosaurier wie
der bis zu 10 Meter lange Plateosaurus,
einer der ersten Sauropoden.

Plateosaurus
Quelle:
Wikimedia Commons File:Sketch plateosaurus.jpg,
von Tim Bekaert (2006), dort gemeinfrei (public domain).
Die Dinosaurier sind in der Trias bereits sehr erfolgreich und verdrängen nach und nach viele Reptilienarten, insbesondere die säugetierähnlichen Reptilien (Therapsiden). Im Jura wird schließlich jedes uns bekannte größere Landtier ein Dinosaurier sein. Insgesamt dominieren Dinosaurier für 150 Millionen Jahre die Erde, bis Sie schließlich 65 Millionen Jahre vor der Gegenwart aussterben. Damit sind Dinosaurier eine der erfolgreichsten Tiergruppen der Erdgeschichte!

Pteranodon,
Quelle: Wikipedia Commons File:Pteranodon hharder.jpg,
Quelle dort: The Wonderful Paleo Art of Heinrich Harder,
Heinrich Harder 1916, copyright abgelaufen.

Ichthyosaurier,
Quelle: Wikipedia Commons File:Ichthyosaur hharder.png,
Quelle dort: The Wonderful Paleo Art of Heinrich Harder,
Heinrich Harder 1916, copyright abgelaufen.
Am Ende der Trias, 200 Millionen Jahre vor der Gegenwart, beginnt Pangäa langsam, erste Risse zu zeigen. Das Auseinanderbrechen Pangäas und die Bildung des Atlantiks sowie des Indischen Ozeans kündigen sich an. Dabei kommt es wieder zu größeren Magmafluten, die sich über das Land ergießen.
Ob diese Magmafluten etwas mit dem Massensterben am Ende der Trias zu tun haben, ist unklar. Bei diesem vierten Massensterben der Erdgeschichte sterben etwa 20% der marinen Familien sowie viele Reptilien und die letzten großen Amphibien aus (siehe z.B. Extinction von John Baez, October 19, 2004, http://math.ucr.edu/home/baez/extinction/ ). Insgesamt ist das Massensterben am Ende der Trias deutlich weniger ausgeprägt als Ende des Perm, aber es eröffnet neue Nischen für die Dinosaurier, die diese freien Plätze im Ökosystem des Jura nutzen werden. Mehr dazu im nächsten Kapitel (Jura).
a) Das Gebiss der Säugetiere
b) Die Evolution der Ohren
Da Zähne zu den am besten erhaltenen fossilen Überresten von Wirbeltieren gehören, werden sie von Paläontologen immer mit besonderem Interesse betrachtet. Außerdem können sie viel über die Lebensweise und die bevorzugte Nahrung eines Tiers verraten.
Säugetiere haben ein besonders komplexes und ausgereiftes Gebiss, das charakteristisch für unterschiedliche Säugetierarten ist. Ihre Zähne besitzen unterschiedliche Höcker, Vertiefungen und Leisten, wobei sich die weiter vorne und weiter hinten sitzenden Zähne stark unterscheiden -- man denke an unsere Schneide- und Backenzähne. Dabei passen die Höcker und Vertiefungen der unteren und oberen Zähne präzise zusammen, was man als Okklusion bezeichnet. Daher können wir so präzise kauen und Nahrung sehr effizient zerkleinern. Reptilien können das nicht! So sehen die messerförmigen Zähne eines Krokodils alle ungefähr gleich aus und unterscheiden sich hauptsächlich nur in ihrer Größe. Außerdem besitzen unterschiedliche Reptilienarten recht ähnliche Zähne.

Diese Komplexität des Säugetier-Gebisses hat allerdings auch einen Preis: Während die Zähne von Reptilien immer
wieder erneuert werden, werden sie bei Säugetieren nur ein einziges Mal ersetzt, nämlich beim
Wechsel vom Milchgebiss zu den erwachsenen Zähnen.
Das präzise Kauen mithilfe gut zusammenpassender Ober- und Unterzähne entstand schrittweise vor rund 225 bis 195 Millionen Jahren (siehe Neil Shubin: Der Fisch in uns, S. 79).
Die säugetierähnlichen Reptilien (Cynodonten) zu Beginn dieser Zeitspanne sahen zwar schon recht hundeähnlich aus, aber ihre Zähne waren noch sehr reptilienähnlich und wurden immer wieder erneuert.
Eine Zwischenstufe zu den Säugetieren sind beispielsweise die Tritheledonten, die ebenfalls zu den Cynodonten gehören. Ihre Zähne besitzen nur eine Wurzel, so wie typische Reptilienzähne, während die Backenzähne von Säugetieren mehrere Wurzeln aufweisen. Andererseits findet man kleine Höcker und Leisten auf den Zähnen, deren typisches Abnutzungsmuster zeigt, dass obere und untere Zähne schon recht gut zusammenpassen -- gleichsam ein Reptil mit präziser Okklusion, wie Shubin es ausdrückt (Neil Shubin: Der Fisch in uns, S. 89). Außerdem unterscheiden sich die vorderen und hinteren Zähne bereits deutlich voneinander.

Vor rund 200 Millionen Jahren gibt es dann mit Morganucodon ein spitzmausartiges Lebewesen,
das den Säugetieren schon sehr ähnlich ist.
Diese Tiere konnten schon präzise kauen, ähnlich wie heutige Säugetiere.
Sie besaßen verschiedene Zahntypen (Schneidezähne, Eckzähne, Prämolare = vordere Backenzähne
und Molare = hintere Backenzähne mit drei Höckern), die nur noch einmal erneuert wurden
und bereits präzise zusammenpassen.

Die Ohren der Säugetiere und damit auch unser eigenes Ohr sind komplexe Gebilde, die im Lauf eines langen Evolutionsprozesses entstanden sind. Das sichtbare Außenohr besteht aus einer Ohrmuschel, die den Schall auffängt und über den Gehörgang bis zum Trommelfell leitet. Die Vibrationen des Trommelfells werden dann im Mittelohr von einer Hebelkette aus drei winzigen Gehörknöchelchen (Hammer, Amboss und Steigbügel genannt) zur sogenannten Schnecke im Innenohr weitergeleitet und über die Membran des ovalen Fensters (auf dem der Fuß des Steigbügels sitzt) auf das Gel übertragen, das sich im Inneren der Schnecke befindet. An der Innenwand der Schnecke befinden sich Sinneszellen mit feinen Härchen, die in das Gel hineinragen. Diese Härchen werden durch die Schwingungen des Gels periodisch verbogen. So können diese Sinneszellen die Vibrationen des Gels spüren und entsprechende Signale an das Gehirn weitergeben.

Neben der Schnecke befinden sich im Innenohr weitere Röhren und Hohlräume, die analog zur Schnecke ebenfalls mit Gel gefüllt und mit feinen Haar-Sinneszellen (kurz Haarzellen) ausgestattet sind. Hier werden allerdings keine Schallvibrationen ermittelt, sondern Informationen über Lage und Beschleunigung des Kopfes gemessen. Mit anderen Worten: hier sitzt unser Gleichgewichtssinn, und er funktioniert nach demselben Grundprinzip wie unser Hörsinn. Es kommt also darauf an, an welchem Ort in den gelgefüllten Hohlräumen des Innenohrs die Gelbewegung von den Haar-Sinneszellen aufgenommen wird, um festzulegen, ob der entsprechende Nervenimpuls als Geräusch, Lage- oder Beschleunigungsinformation wahrgenommen wird. Die Funktion als Gleichgewichtsorgan ist dabei sehr viel älter als der Gehörsinn -- das Innenohr hat also erst deutlich später im Lauf der Evolution auch hören gelernt.
Wozu brauchen Säugetiere diese komplexe Hebelkette aus den drei Gehörknöchelchen? Könnte der Schall nicht unmittelbar aus der Luft in das Gel der Schnecke eintreten und dieses in Schwingungen versetzen?
Schall tritt nur sehr ungern von einem wenig dichten, gut komprimierbaren Medium wie Luft in das flüssige, viel dichtere und kaum komprimierbare Gel über. Der meiste Schall würde also reflektiert, ähnlich wie Licht von einer Wasseroberfläche reflektiert wird, und würde nicht die feinen Haar-Sinneszellen im Schneckeninneren erreichen.
Schall führt in der Luft zu recht großen Schwingungsamplituden, aber nur kleinen Druckamplituden. Die Luftmoleküle schwingen also relativ weit, aber nur mit wenig Kraft. In Flüssigkeiten entstehen dagegen nur kleine Schwingungsamplituden, aber große Druckamplituden, da Flüssigkeiten viel weniger stark verdichtet (komprimiert) werden können als Gase -- Flüssigkeiten sind nahezu inkompressibel. Daher bewegen sich die Flüssigkeitsmoleküle beim Durchgang von Schallwellen nur relativ wenig, dafür aber mit großer Kraft.
Das Hebelsystem der Gehörknöchelchen wandelt nun große Schwingungsamplituden und kleine Druckamplituden in kleine Schwingungsamplituden und große Druckamplituden um, sodass die Vibrationen sich gut auf das Gel im Inneren der Schnecke übertragen lassen. Das Hebelsystem der Gehörknöchelchen wirkt also als physikalischer Impedanzwandler und ermöglicht es uns erst, leise oder besonders hohe Töne noch wahrzunehmen.
Das Gehör von Säugetieren ist einmalig: Nur sie besitzen eine äußere Ohrmuschel und ein Hebelsystem aus drei Gehörknöchelchen. Die drei Gehörknöchelchen sind ähnlich wie die spezielle Form der Zähne (siehe oben) ein charakteristisches Kennzeichen der Säugetiere, das sich gut in Fossilien nachweisen lässt (anders als die Existenz von Milchdrüsen).
Bleibt die Frage, wie ein so komplexes System wie unser Ohr im Lauf der Evolution überhaupt entstehen konnte.
Das Gehör von Amphibien, Reptilien und Vögeln ist deutlich anders aufgebaut. Sie besitzen nur einen einzigen Gehörknochen: den Steigbügel. Dieser hat allerdings bei ihnen nicht die typische Steigbügelform wie bei den Säugetieren, sondern ist ein längliches Knochenstäbchen, das auch als Säulchen (Columella) bezeichnet wird. Das eine Ende dieses Säulchens ist mit dem einfacher genbauten Innenohr verbunden, das andere Ende mit dem Trommelfell oder direkt mit dem Schädelknochen -- im zweiten Fall werden keine Luftschwingungen erfasst, sondern Bodenvibrationen.
Wie konnten die beiden zusätzlichen Gehörknöchelchen entstehen? Man geht heute davon aus, dass Hammer und Amboss aus zwei Knochen am hinteren Ende des Kiefers der Reptilien hervorgegangen sind. Bei Reptilien bilden sie als Articulare und Quadratum das (primäre) Kiefergelenk, das bei Säugetieren dem Gelenk zwischen Hammer und Amboss entspricht. Dafür haben Säugetiere an etwas anderer Stelle ein anderes (sekundäres Kiefergelenk).
Man kennt heute viele fossile Übergangsformen zwischen säugetierähnlichen Reptilien und den ersten Säugetieren, an denen sich die Evolution der Gehörknöchelchen gut nachvollziehen lässt. Dabei wurden die Knochen am hinteren Ende des Kiefers immer kleiner und wanderten ins Mittelohr. Beim Menschen wiegt der Hammer schließlich nur noch 23 mg, der Amboss 27 mg und der winzige Steigbügel sogar nur 2,5 mg. Hammer und Amboss wurden fossil zuerst bei Hadrocodium wui (früher Jura) nachgewiesen.
Den unterschiedlichen Ursprung der drei Gehörknöchelchen kann man auch an der embryonalen Entwicklung ablesen: Hammer und Amboss stammen vom ersten Kiemenbogen ab, genau wie der Kiefer. Der winzige Steigbügel stammt dagegen vom zweiten Kiemenbogen ab (zum Thema Kiemenbögen siehe auch Kapitel 4.1 Zusatzinformationen).
Es scheint nicht ganz unumstritten zu sein, dass die ersten amphibienartigen Tetrapoden gleichsam das Gehör mit dem Steigbügelknochen zuerst entwickelt und dann an die Reptilien und schließlich die Säugetiere vererbt haben, wobei die Säugetiere noch zwei weitere Knochenteile nachträglich aus den Kiefern integriert haben. Eine solche nachträgliche Integration weiterer Knochen in die Hebelkette des Mittelohrs wirft Probleme auf, da ja jedes Zwischenstadium voll funktionsfähig sein muss. Möglich ist auch, dass Amphibien, Reptilien und Säugetiere erst nach einiger Zeit unabhängig voneinander und fast gleichzeitig entsprechende Mittelohrknochen entwickelt haben (siehe Benedikt Grothe: Wie die Evolution die Ohren aufsperrt, http://www.mpg.de/1043263/F001_Fokus_024_031.pdf). Die Wege der Evolution sind komplex, und so manche Lehrmeinung mag sich ändern, sobald weitere Details ans Licht kommen.
Gehen wir noch weiter in unserem Stammbaum bis zu den Fischen zurück, so findet man gar keine Gehörknöchelchen mehr. Dafür findet man andere Knochen, insbesondere bei Haien und Knochenfischen die Hyomandibula, die als Knochenstab den Oberkiefer beweglich am Schädel aufhängt -- deshalb können Haie beim Zubeißen ihren Oberkiefer so charakteristisch aufklappen.
Bei der Entwicklung zu den Tetrapoden leitete dieser Knochen möglicherweise zunächst Bodenvibrationen weiter und könnte so zum Vorläufer des Steigbügels geworden sein, wobei er nach und nach immer kleiner wurde, sodass er zunehmend für seine neue Funktion als Vibrations-Weiterleitung optimiert wurde. Bei Säugetieren wurde der Steigbügel dann schließlich noch kleiner und nahm seine typische Steigbügel-Form an.
Interessanterweise ist auch bei vielen Fischen unabhängig ein eigenes Gehör entstanden. Dazu muss man wissen, dass einfache Fische akustisch zunächst weitgehend transparent sind, d.h. eine Schallwelle würde bei ihnen die flüssigkeitsgefüllten Haar-Sinneszellen synchron mit der sonstigen Flüssigkeit mitbewegen und zu keiner Auslenkung der feinen Haare führen. Das ändert sich bei Fischen, die eine Schwimmblase besitzen, denn in der luftgefüllten Schwimmblase breitet sich Schall wesentlich langsamer aus als in Flüssigkeit, sodass man hier ähnlich wie bei einem Trommelfell Schallvibrationen abgreifen kann. Die Weiterleitung zum Labyrinth des Innenohrs geschieht hier durch die Weberschen Knöchelchen (Weberscher Apparat), die Abkömmlinge von Rippen der Halswirbel sind.
Wie schon bei der Evolution der Augen (siehe Kapitel 4.1 Zusatzinformationen) sehen wir, dass auch ein Gehör auf unterschiedliche Arten mehrfach im Lauf der Evolution enstehen kann. Sogar viele Insekten können Schall wahrnehmen, wenn auch auf eine ganz andere Weise als Wirbeltiere -- sonst würden Grillen nicht zirpen, um einen Fortpflanzungspartner anzulocken (siehe Zeit online: Hör zu, Fliege!, http://www.zeit.de/2008/16/P-Goepfert).
Analog zum Auge hat auch das das Gehör der verschiedenen Tiere eine gemeinsame molekulare und zelluläre Basis. Diese gemeinsame Basis bilden die Haar-Sinneszellen. Schon die ersten kieferlosen Fische besaßen solche Haarzellen, und auch das heutige Lanzettfischchen hat sie, obwohl es weder Augen noch Ohren oder einen Schädel besitzt. Sie finden sich bei Fischen im Seitenlinienorgan (in den sogenannten Neuromasten), mit dem sie die Strömungsverhältnisse im umgebenden Wasser sehr genau wahrnehmen können. Tatsächlich entstehen im Embryo Innenohr und Neuromasten aus demselben Gewebe, wobei nicht klar ist, welches von beiden entwicklungsgeschichtlich zuerst da war.
Ähnlich wie beim Augen-Hauptschalter-Gen Pax 6 gibt es auch für das Gehör eine allgemeine genetische Basis: So ist das Gen Pax 2 bei Säugetieren für Entwicklung des Innenohrs wichtig und bei Fischen für die Entwicklung des Seitenlinienorgans.
Interessanterweise haben das Augen-Gen Pax 6 und das Gehör-Gen Pax 2 womöglich sogar einen gemeinsamen Ursprung in den Tiefen der Evolution: Das Augen-Gen der Würfelqualle (die übrigens sogar einfache Linsenaugen besitzt) sieht aus wie eine vereinfachte Mischung der Gene Pax 6 und Pax 2. Würfelquallen gehören als Nesseltiere zu einem Zweig der Evolution, der sich noch vor der Entstehung unseres eigenen Zweiges der Zweiseitentiere (Bilateria) selbstständig gemacht hat.
Literatur zu dem Thema:
last modified on 30 June 2012
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}